蛋白质分选枢纽
反面分选
糙面内质网合成多种蛋白,而高尔基体的 TGN 区则负责蛋白质包装分选,有以下三种途径
- 溶酶体酶的包装分选途径 ^9f7839
- 溶酶体酶在高尔基体经过一系列修饰带上 M6P (6-磷酸甘露糖)标记
- 在高尔基体TGN区的膜上,存在M6P受体。带有M6P标记的溶酶体酶与这些受体特异性结合。通常一个溶酶体酶有多个M6P位点,这大大增强了与受体的亲和力,从而将溶酶体酶从众多蛋白质中“筛选”出来并局部浓缩。
- 溶酶体酶-M6P受体复合物通过出芽的方式,形成由衔接蛋白包被的运输小泡,离开高尔基体
- 运输小泡与晚期内体(Late Endosome) 融合,晚期内体内的酸性环境导致溶酶体酶与M6P受体分离
- 调节型分泌:如促肾上腺皮质激素(ACTH)、胰岛素等
- 组成型分泌:如流感病毒核水泡性口炎,说明分选信息可能存在于蛋白质自身
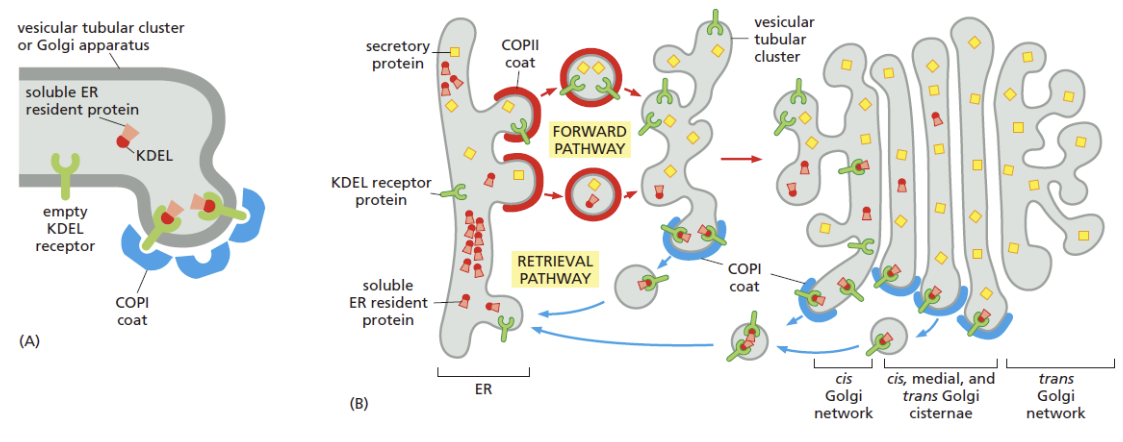
顺面分选
除反面,顺面也对蛋白质的分选和运输,帮助内质网实现内质网蛋白质的选择性分泌和维持
- COPⅡ、COPⅠ和高尔基体CGN 区膜上均有识别与结合KDEL 信号(四肽驻留信号)的受体
- CGN 区的 KDEL 信号受体可识别高尔基体内的内质网蛋白,募集 COP I 包被囊泡,并将其返还至 ER
- ER 与高尔基体内 pH 不同,促进的驻留蛋白分选信号和受体的结合和释放
蛋白质糖基化
他妈的,太烦了,我觉得前面写过了
糖脂的合成
糖脂的糖侧链以与糖蛋白相同的途径和方式在高尔基体合成与加工,由高尔基体转运到溶酶体膜或细胞膜上
植物细胞壁的合成
高尔基体合成和分泌的多糖参与细胞壁的构成,虽然纤维素是由细胞膜外侧的纤维素合成酶合成的
蛋白酶的水解和其他加工过程
在高尔基体的 TGN 甚至 TGN 所形成的分泌小泡中时,部分肽链经蛋白水解酶的特异性水解才成为有生物活性的多肽
- 没有生物活性的蛋白原(proprotein)进入高尔基体后将蛋白原的N端或两端的序列切除形成成熟的多肽
- 有些蛋白质在粗面内质网中合成时是含有多个相同氨基酸序列的前体,然后在高尔基体中水解成同种有活性的多肽
- 一个蛋白分子的前体中含有不同的信号序列,最后加工成不同的产物。增加了细胞信号分子的多样性
硫酸化作用也在高尔基体中进行(硫酸化的蛋白质主要是蛋白聚糖)